
通讯:13983250545
微信:ycsh638
QQ:469764481邮箱:ycsh6318@163.com
论文:温度诱导鳜三倍体的比较研究
温度诱导鳜三倍体的比较研究
魏 晋,宋 文,王守荣,陈元元,曾可为
(1.武汉市农业科学院,武汉 430065;2.江汉大学,武汉 430056)
鳜(Sinipercachuatsi)隶属于鲈形目(Perciformes)鲈亚目(Percoidei)真鲈科(Percichthyidae)鳜属(Siniperca),在中国、俄罗斯、日本、韩国、朝鲜以及越南等国均有分布,在我国的中东部平原区分布最为广泛。鳜与其他鱼类相似,在性腺发育阶段及产卵季节会消耗更多的能量,存在生长减缓、抗逆能力下降及肉质变差等现象。采用现代生物学手段,开发鳜的三倍体诱导技术,可以有效提升鳜的生长速度和抗逆能力,降低其饵料系数,对缩短鳜的养殖周期,提升产量和肉质,提高养殖经济效益,具有重要意义和实际价值[1]。
人工诱导三倍体鱼形成已先后在40多种海水和淡水鱼类中成功,其中三倍体兴国红鲤、湘云鲫、湘云鲤、异育银鲫和三倍体虹鳟等品种已进入规模化生产阶段,带来了巨大的经济效益[2,3]。本研究参考了泥鳅(Misgurnusanguillicaudatus)[4]、黄颡鱼(Pelteobagrusfulvidraco)[5,6]、团头鲂(Megalobramaamblycephala)[7]、乌鳢(Channaargus)[8]、细鳞鲑(Brachymystaxlenok)[9]及鳜(S.chuatsi)[10]等淡水鱼类的三倍体诱导方法,采用冷休克和热休克法进行了鳜三倍体的诱导研究,筛选到了最佳的诱导条件,对冷、热休克法诱导的效果进行了综合比较,并对三倍体和二倍体鳜鱼苗的生长性状及存活率进行了研究,以期为进一步开展鳜三倍体苗种的规模化生产提供理论基础和实践依据。
1 材料与方法
1.1 亲本来源及受精卵的获取
本试验所采用的鳜亲本采集自武汉市佳恒水产有限公司养殖基地。在鳜的繁殖季节,挑选表型正常、无伤病、身体健壮并且已达性成熟的2龄雌鱼和雄鱼各30尾进行人工繁殖,雌鱼体重750~950 g,雄鱼体重650~800 g。
采用人工催产的方法获得鳜受精卵,产前培育、药物催产及孵化管理等具体步骤参照李志成等[11]和刘希良等[12]的方法。
1.2 温度休克处理最佳诱导条件的筛选
本研究中,在鱼苗发育不同阶段,统计各处理组和对照组的受精率、存活率、畸形率和三倍化率。受精率(FR):胚胎发育原肠胚期,存活胚胎数目占总卵数目的比例。存活率:孵化后5 d,存活个体数目占受精卵数目的比例;相对存活率(SR)=处理组存活率/对照组存活率×100%。畸形率:孵化后5 d,畸形个体数目占存活个体数目的比例;相对畸形率(MR)=处理组畸形率/对照组畸形率×100%。三倍化率(TR):孵化后8 d从各处理组随机挑选50尾鱼苗检测其倍性,其中三倍体鱼苗所占比例。数据处理软件为SPSS 13.0和Microsoft Office Excel 2003。
1.2.1 冷休克处理最佳诱导条件的筛选
冷休克试验过程参照宋立民等[5]的方法,采用单因子试验获得最适的冷休克诱导条件。三个单因素分别为:处理起始时间(BT)、处理温度(Temp)和处理持续时间(DT)。试验设置如表1,各组经冷休克处理后,受精卵转移至26 ℃水体中继续培养。设置对照组:受精卵始终置于26 ℃水温条件下培养。每个处理组和对照组均设置3个重复,每个重复的受精卵数量约2 000粒。
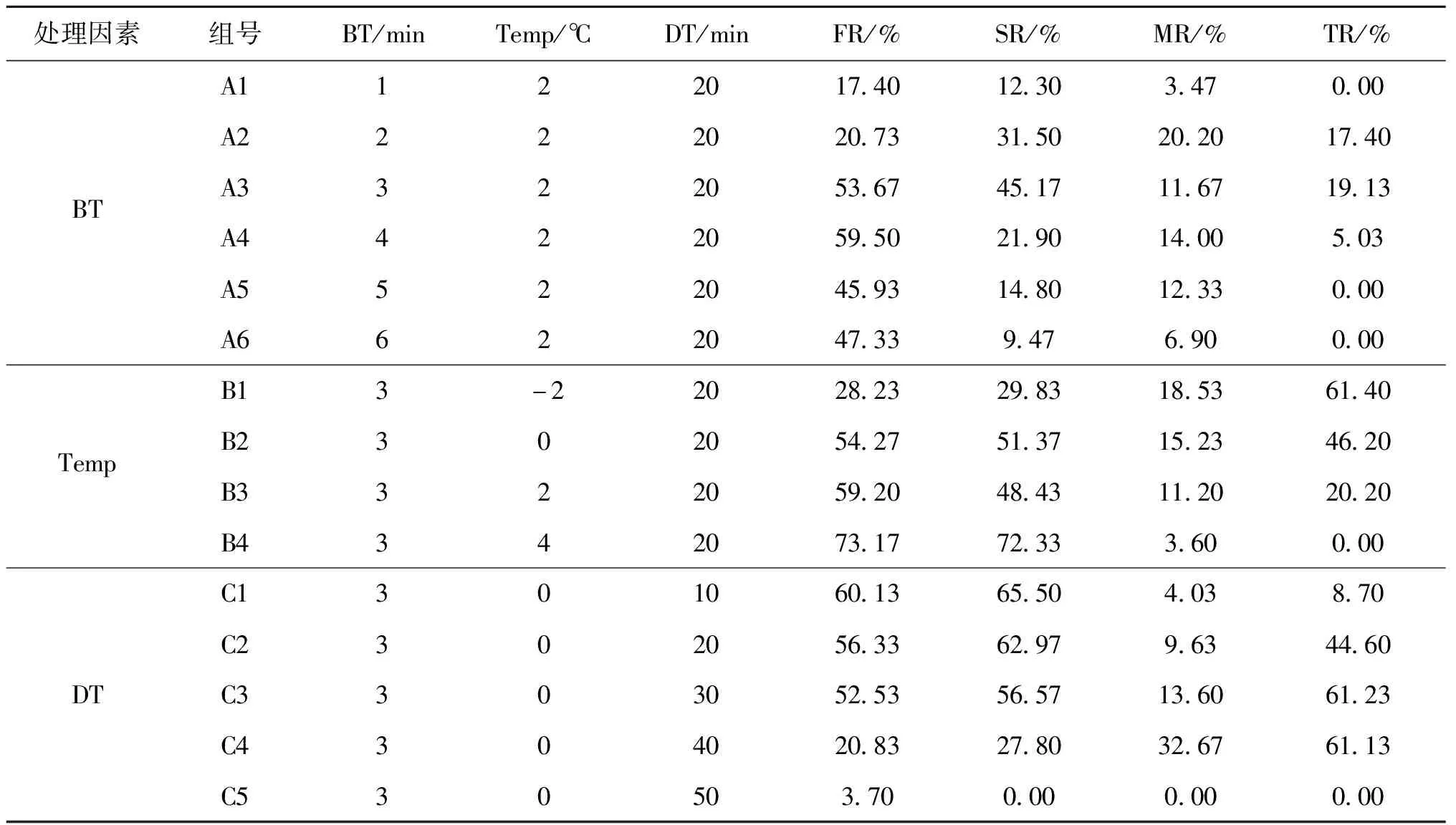
表1 冷休克单因素处理试验设置
1.2.2 热休克处理最佳诱导条件的筛选
鱼类三倍体的热休克诱导试验一般于亚致死温度附近实施,参考Xu等[10]的研究结果进行预实验,发现当水温高于40 ℃后,鱼苗存活率为0%,因此,本研究将热休克处理温度范围设置为32~38 ℃。其他单因素试验条件设置如表2,各组经热休克处理后,受精卵转移至26 ℃水体中继续培养。设置对照组:受精卵始终置于26 ℃水温条件下培养。每个处理组和对照组均设置3个重复,每个重复的受精卵数量约为2 000粒。
![]()
表2 热休克单因素处理试验设置
1.3 倍性检测
孵化后8 d的鳜鱼苗体型较小,无法进行尾柄采血,因此从每组中随机挑选50尾鱼苗,制备单细胞悬液进行倍性检测。将每尾待检鳜鱼苗放入2 mL EP管中,蒸馏水清洗2次后,加入0.4 mL PBS(pH 7.4)溶液,利用组织破碎仪将鱼苗碾碎,经300目筛绢过滤后加入PBS(pH 7.4)溶液制备成细胞浓度约为103个/mL的单细胞悬液。1月龄以后的幼鱼,通过尾部静脉取血,利用PBS(pH 7.4)溶液制备成浓度约为103个/mL的血细胞悬液进行倍性检测。
制备好的细胞悬液采用4′,6-二脒基-2-苯基吲哚(4′,6-diamidino-2-phenylindole,DAPI)染色 1 min,用流式细胞仪(Beckman Coulter,CytoFlex,USA)进行 DNA 含量分析。采用正常二倍体鳜鱼苗作为对照。本研究中二倍体鳜细胞DNA相对含量为(204.2±4.3),三倍体鳜细胞的DNA相对含量为(301.1±6.1),约为二倍体的1.47倍(图1),可判断诱导出的鳜鱼苗为三倍体。
![]()
图1 二倍体(a)和三倍体鳜(b)细胞 DNA 相对含量直方图
1.4 冷、热休克诱导效果比较及鳜三倍体鱼苗生长性能检测
为排除亲本遗传差异对试验结果的影响,在确定冷、热休克法的最优诱导条件后,利用3对鳜亲本分别构建3个全同胞家系,对两种方法的诱导效果进行比较。每个家系随机选取约2 000颗受精卵,采用优化后的冷、热休克诱导条件分别各处理1 000颗以获得三倍体鱼苗;剩余受精卵在正常条件下培育,以获得二倍体鱼苗作为对照组。
所有鱼苗均在同等适宜条件下培育,充足投饲,分别在孵化后10、20、40、60、100和150 d,从温度休克处理组和正常对照组中随机抽取50尾幼鱼进行倍性鉴定,并测量其体长和体重。剩余鱼苗继续培养至180 d(幼鱼期),统计三个家系中,采用冷、热休克法所产生后代的存活率及TR。三倍体和二倍体鳜在不同发育阶段生长差异分析采用t检验,统计分析及作图均利用SPSS 13.0软件完成。统计值采用平均值±标准差(Mean±SD)表示,P<0.05表示具有显著差异,P<0.01表示具有极显著差异。
2 结果与分析
2.1 冷休克最佳诱导条件的确定
在设定条件(Temp=2 ℃,DT=20 min)下进行冷休克处理最佳起始时间的筛选,结果表明受精卵在受精后2、3和4 min进行冷休克处理都能诱导产生三倍体鳜鱼苗,其中受精后3 min进行冷休克处理的SR和TR均达到峰值(表1,图2A),因此采用冷休克法诱导鳜三倍体的最佳起始时间为受精后3 min。在设定条件(BT=3 min,DT=20 min)下进行冷休克处理最适温度的筛选,结果表明在-2、0和2 ℃下,均可以诱导产生三倍体鳜鱼苗,其中在-2 ℃下处理获得后代的TR虽然最高,但FR和SR相对较低;而在0 ℃条件下处理获得的后代,其FR、SR和TR均相对较高(表1,图2B),因此综合考虑,采用冷休克法诱导鳜三倍体的最适处理温度为0 ℃。在设定条件(BT=3 min,Temp=0 ℃)下进行冷休克处理持续时间的筛选,结果表明冷休克处理10、20、30和40 min都能诱导产生三倍体鳜鱼苗,其中处理30 min和40 min时TR相对更高,分别可达到61.2 %和61.1 %,但是冷休克处理40 min的FR和SR急剧下降,而MR则达到峰值(表1,图2C),因此综合考虑,采用冷休克法诱导鳜三倍体的最适处理持续时间为30 min。
![]()
图2 不同冷休克诱导条件下FR、SR、MR及TR的变化
2.2 热休克最佳诱导条件的确定
在设定条件(Temp=36 ℃,DT=2 min)下进行热休克处理最佳起始时间的筛选,结果表明受精卵在受精后6、7和8 min进行诱导可产生三倍体鳜鱼苗,并且FR、SR和MR无显著差异(P>0.05),其中受精后7 min进行热休克处理的TR达到峰值,平均可达到26.6%(表2,图3A),因此采用热休克法诱导鳜三倍体的最佳起始时间为受精后7 min。在设定条件(BT=7 min,DT=2 min)下进行热休克处理最适温度的筛选,结果表明在34、36和38 ℃下诱导均能产生三倍体鳜鱼苗,并且FR、SR和MR无显著性差异,其中38 ℃下诱导的TR达到峰值,平均可达到30.0%(表2,图3B),因此综合考虑,采用热休克法诱导鳜三倍体的最适处理温度为38 ℃。在设定条件(BT=7 min,Temp=38 ℃)下进行热休克处理持续时间的筛选,结果表明热休克处理1 min、2 min和3 min均能诱导产生三倍体,其中处理3 min后TR达到峰值,尽管FR和SR略低于2 min时,但并无显著性差异(表2,图3C),因此综合考虑,采用热休克法诱导鳜三倍体的最适处理持续时间为3 min。
![]()
图3 不同热休克诱导条件下FR、SR、MR及TR的变化
2.3 优化条件下冷、热休克方法的比较
由于亲本的遗传差异也会对温度休克的效果造成影响[10],因此本研究在获得两种方法的优化条件后,又构建了三个全同胞家系来比较冷、热休克法诱导三倍体鳜的实际效果。结果表明,采用冷休克法诱导(BT=3 min,Temp=0 ℃,DT=20 min),幼鱼期(180 dph)平均存活率约为40.9%,其中平均TR约为55.40%;采用热休克法诱导(BT=7 min,Temp=38 ℃,DT=3 min),三个家系均在孵化后25~30 d开始出现大量发育畸形的情况,最终1号和2号家系的TR尽管分别可达到26.97% 和22.81%,但幼鱼期存活率均未超过10%,而3号家系幼鱼期存活率仅为1.6%,其中无三倍体幼鱼(表3)。该结果表明冷休克诱导更适用于鳜三倍体鱼苗的规模化生产。
![]()
表3 冷、热休克方法诱导效果比较
2.5 二倍体和三倍体鳜鱼苗的生长性能比较
饲养过程中观察到,二倍体和三倍体鱼苗在卵黄囊吸收完全后均出现死亡高峰,而三倍体群体在体长约为20 mm时又出现一个明显的死亡高峰,其中多数死亡个体为畸形,在饲养过程中未出现明显的发病现象。对不同发育阶段的二倍体和三倍体鳜的体长和体重进行t检验,结果表明孵化后10、20和40 d的二倍体与三倍体鳜鱼苗的体长和体重均无显著差异,60、100和150 d的三倍体鳜鱼苗体长显著高于二倍体,体重极显著高于二倍体。生长速率方面,在孵化后40 d内,二倍体和三倍体鳜的体长和体重平均增长率均无显著差异;孵化后40~150 d,二倍体的体长和体重平均增长率分别为0.102 cm/d和3.414 g/d,三倍体的体长和体重平均增长率分别为0.159 cm/d和5.037 g/d,均显著高于二倍体;其中在孵化后40~60 d阶段,二倍体和三倍体的生长速率均达到最高(表4,图4)。综上所述,鳜三倍体鱼苗的生长性能优于二倍体,但也存在畸形率较高的问题。
![]()
表4 二倍体和三倍体鳜在不同发育阶段体长和体重比较
![]()
图4 二倍体和三倍体鳜生长性能比较
3 讨论
3.1 冷、热休克诱导效果的比较
人工诱导鱼类三倍体的主要方法包括冷休克法[13]、热休克法[14]以及静水压法[15]。由于不同鱼类的遗传背景、生物学特性以及对温度或压力的敏感性有差异,因此这些方法不一定适用于所有的鱼类,并且不同方法的诱导效率通常也有区别。静水压法的诱导率通常高于温度休克法。例如在人工诱导大黄鱼(Pseudosciaenacrocea)三倍体时发现,静水压法的诱导率要明显优于冷休克法[16]。不过由于静水压休克设备要求较高,限制了批量化生产的效率,因此目前的研究更多采用温度休克法诱导鱼类三倍体。选择冷休克法还是热休克法需要根据不同物种自然生活环境的水温来判断,并通过反复试验确定[17]。因为不同鱼类受精卵对温度的敏感性不同,从而造成了耐受温度的差异,通常温水性鱼类采用冷休克法诱导,而冷水性鱼类常采用热休克法诱导,冷、热刺激的温度均不能超过该物种的自然致死温度[18]。
Xu等[10]采用热休克法诱导鳜三倍体时发现,在受精后8 min将受精卵置于41 ℃下热休克处理2 min,可获得40%的三倍体检出率。本研究的结果与之有所区别,当孵化水温高于40 ℃后,发现鳜受精卵及鱼苗几乎无法存活,这一结果与何利君[19]的结论较为一致。而在优化后的热休克处理条件下,尽管仔鱼期(孵化后8 d)的三倍体检出率可达到38.0%,但在孵化后25~30 d开始出现大量发育畸形的情况,最终三倍体幼鱼的存活率不足3%,推测可能是鳜的胚胎发育对高温较为敏感导致的[19]。此外,在优化后的冷休克处理条件下,仔鱼期(孵化后8 d)和幼鱼期的三倍体检出率分别为61.2% 和56.0%,表明相比于热休克诱导,冷休克诱导更适用于鳜三倍体的规模化生产。不过本研究在饲养冷休克诱导的三倍体鳜时也发现,鱼苗在体长约为20 mm时出现了一个明显的死亡高峰,其中多数死亡个体为畸形,这可能是由于冷休克处理的时间较长,对胚胎造成了一定损害所导致的[18]。宋立民等在采用冷、热休克两种方法诱导黄颡鱼三倍体时也发现了类似的现象[5]。
3.2 影响温度休克诱导效果的因素
结合国内外学者对多种鱼类进行温度休克诱导的研究结果,适宜的休克起始时间、休克温度和持续时间是保证三倍体诱导成功的关键因素和重要前提。而在不同鱼类中或者在同一物种中利用不同的温度休克方法时,这三个关键因素的重要性主次顺序也有所不同。尤锋[20]在采用冷、热休克法对黑鲷(Sparusmicrocephalus)进行三倍体人工诱导时发现,两种方法的三倍体率受三个因素影响程度的顺序均依次为:持续时间>起始时间>处理温度。而利用冷休克法诱导红鳍东方鲀(Takifugurubripe)三倍体[21]和利用热休克法诱导虹鳟(Oncorhynchusmykiss)三倍体[22]时,温度休克持续时间也是影响三倍体率的最重要因素。宋立民等[5]利用冷休克法对黄颡鱼进行三倍体人工诱导时发现,休克起始时间是TR和孵化率的重要影响因子,处理温度则是畸形率的重要影响因子;而利用热休克法诱导三倍体时,处理温度是三倍体率、畸形率和SR的重要影响因子,次之为起始时间和持续时间。产生这种差异的原因目前尚不明确,而除了上述三个重要因子外,不同物种当中影响多倍体诱导效果的其他因素也不尽相同,亲本的遗传差异,受精卵的质量和生物性状,以及休克处理时的具体环境条件等都是值得考虑的因素[5,9]。目前在关于鳜鱼三倍体诱导的研究中,尚无对多因素影响的讨论[10],未来将对此进行更为深入的研究。
3.3 二倍体和三倍体鳜鱼苗的生长性能比较
人工诱导鱼类三倍体的主要目的,是希望通过阻碍其性腺发育,提高生长速度和抗逆性能,提升肉质和产量,此外由于三倍体的不育性,也能有效防止其对原始生态系统的破坏[1]。然而关于三倍体鱼类生长性能的研究,不同品种的鱼类其结果也有差异。例如,杨兴棋和陈敏容利用二倍体白鲫(Carassiusauratus)与四倍体白鲫交配获得了异源三倍体白鲫,相比正常二倍体,其体型更大,生长更快[23]。而在欧洲黑鲈(Dicentrarchuslabrax)[24]和乌鳢(C.argus)[8]中发现,三倍体与二倍体的生长速度无显著差异。大菱鲆(Scophthalmusmaximus)三倍体在性成熟前,与二倍体的生长无显著差异,但达到初次性成熟后,三倍体生长显著快于二倍体[25]。本研究发现,孵化后10~40 d的三倍体与二倍体鳜的生长速度无显著差异,孵化后60~150 d的三倍体生长速度显著快于二倍体,随着二倍体鳜性腺的发育,两者的生长性能变化仍需进一步研究。
4 结论
综上所述,受精后3 min将受精卵置于0 ℃下冷休克处理20 min,然后转移至26 ℃水体中继续培养,此方法获得鱼苗的受精率为52.5%,相对存活率为56.6%,相对畸形率为13.6%,三倍化率为61.2%;最优的热休克诱导条件为:受精后7 min将受精卵置于38 ℃下热休克处理3 min,然后转移至26 ℃水体中继续培养,此方法获得鱼苗的受精率为30.4%,相对存活率为33.4%,相对畸形率为14.5%,三倍化率为38.0%。通过比较冷、热休克法诱导的鱼苗在幼鱼期的存活率和三倍化率,证明了冷休克诱导更适用于鳜三倍体鱼苗的规模化生产。通过比较二倍体和三倍体鳜在不同生长阶段的生长速率,证明了三倍体鳜鱼苗具有更为优良的生长性能。后续需要对冷休克法诱导鳜三倍体的条件进一步优化,提高诱导率,降低畸形率,以期达到满足规模化生产的要求。
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!



