
通讯:13983250545
微信:ycsh638
QQ:469764481邮箱:ycsh6318@163.com
论文:不同运动模式下鲢幼鱼游泳特性研究
不同运动模式下鲢幼鱼游泳特性研究
郝文超,王从锋,2,刘慧杰,王 瑶,冯三杰
(1.三峡大学水利与环境学院,湖北宜昌 443002;2.三峡地区地质灾害与生态环境湖北省协同创新中心,湖北宜昌 443002)
鱼类作为水生生态系统信息和能量循环正常运行的关键因子,相关研究表明水利工程建设切断河流的连通性,阻隔鱼类洄游通道,对鱼类生境造成严重的破坏,进而导致鱼类资源急剧减少[1]。近年来,通过人工增殖放流[2]和鱼类栖息地适宜度评估手段[3]对珍稀鱼类及特有物种进行保护取得一定进展,但由于保护物种自身的局限性和自然条件的不可控因素,未能从根本上解决问题。修建过鱼设施作为保护鱼类资源的另一重要手段受到研究人员的广泛关注[4]。随着渔业生产的发展和现代技术的进步,鱼类行为学研究逐渐开展起来,为水利工程过鱼、拦鱼和诱鱼等设施提供必不可少的基础数据[5-7],其中目标鱼类游泳能力及其能量代谢特征是过鱼设施修建的基本原则[8]。
目前国内外为了获得鱼类游泳稳定的线性运动,科研人员多数会采用特定的实验装置,进行鱼类强迫式游泳的实验,来研究鱼类游泳特性[9,10],而在自然状态下,鱼类的游泳多以自发性游泳方式进行,与水环境生态因子变化或其自身的生理需求,如摄食、繁殖等行为相关。鱼道设计过程中,需要更为贴近自然情况下鱼类游泳特性的参考依据,在实际情况下,还不能直接观测自发运动条件下的运动消耗,但可以通过与耗氧率Mo2相关的鱼类游泳特性进行表征[11,12],所以自发运动条件下游泳特性分析显得尤为重要。
四大家鱼是中国典型江湖洄游性鱼类,大多数过鱼设施的过鱼对象都包括四大家鱼,国内外关于鱼类游泳能力及其能量代谢特征研究集中在强迫式运动条件下游泳速度及外界条件对其造成的影响[13,14],针对自发性运动的研究尚未见报道。本实验以鲢幼鱼为对象,旨在分析不同运动模式对游泳呼吸代谢的影响,通过对自发性运动的耗氧率与影响因素进行拟合分析,定量的描述两种运动模式下的差异性,为鱼道设计及鱼类行为学研究提供参考。
1 材料与方法
1.1 实验材料
实验鱼来源于湖北宜都水产养殖渔场,鲢幼鱼体长为11.19~12.02 cm,体重为20.84~24.36 g,共24尾。暂养在水温为(25±1)℃的循环控温水槽内,时间14 d。光照条件为16 h白天∶8 h黑夜。每天投足饱饵料1次。实验前转移至2.5 L自来水箱适应2 d,适应条件与暂养条件相同,每天早晚换水1次,每次换水量为20%,实验前禁食2 d。
1.2 实验装置
自发性运动模式测定采用自主研发的船闸式水槽鱼类呼吸装置[15]。船闸式水槽鱼类游泳呼吸装置密闭试验区体积为36 L,长60 cm,宽40 cm,高15 cm。水槽内设置有坝体和船闸,整个装置密封。装置外部上方架设摄像头。实验开始时,启动水泵,打开并调节阀门控制水流速度、冲水角度和流动状态,使形成贴近于自然状态下的复杂流区。采用AutoRespTM软件和HACHHQ30d便捷式水质多参分析仪,分别自动控制流速和瞬时测定温度及溶解氧。设定摄像机视频拍摄频速为21帧/s,采用LoliTrack软件进行视频分析,以获取游泳速度U、旋转角度θ、加速度ΔU和摆尾频率TBF。
强迫式运动模式游泳速度测定采用丹麦Loligo System公司生产的鱼类行为视频跟踪系统(Loligo System swim-large 90 L,PA10300)[16]。实验开始前通过哈希水质多参分析仪对温度以及溶氧进行校核,装置上方架设21帧/s的摄像头连接uEye Cockpit程序和LoliTrack软件进行视频分析,获得摆尾频率TBF数据。
1.3 研究方法
实验水温控制为(25±1)℃,选取健康无病害的鲢幼鱼分为2组,每组12尾,分别进行自发性运动和强迫式运动条件下的实验分析。
1.3.1 游泳速度和耗氧率的测定
自发性运动模式是在船闸式水槽鱼类游泳呼吸装置中进行,每条实验鱼进行8 h观测,采用21帧/s的摄像头进行视频录制,后期数据处理选取60 s时长为一个数据处理单元,每5 min作为一个测量指标的单元,该单元取5 min内测量数据的平均值。通过LoliTrack软件,计算该时长内的游泳速度U、旋转角度θ、加速度ΔU。耗氧率Mo2通过AutoRespTM软件读取,每隔150 s读取一次,5 min内读取2个,结果取平均值。
强迫式运动模式将其放入环形水槽以0.5 BL·s-1[BL为体长(cm)]流速适应1 h,随后以每30 min增加0.5 BL·s-1的梯度进行试验,采用Auto Resp TM软件监测溶解氧,每隔150 s进行溶解氧计数共12次,计算对应其游泳速度下的耗氧率Mo2,直到实验鱼贴网时间超过20 s,视为疲劳。实验设置空白对照,发现无鱼状态下实验过程的耗氧率不超过鱼类耗氧率的1%。计算公式[16]如下:
(1)
(2)
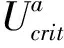
(3)
![]()
对于两种运动模式下的实验,耗氧率Mo2通过游泳速度(U)的幂函数来描述[17],拟合方程如下:
Mo2=a+bUc(4)
式中a,b,c为常数,其中a为标准代谢率SMR。
1.3.2 运动行为学分析
自发性运动模式,在每5 min内选取2个60 s时长的视频。强迫式运动模式,每个游泳速度下选取5个时长为60 s的视频。分别通过LoliTrack软件进行计数取平均值。拟合方程[18]如下:
TBF=a+bU
(5)
lnMo2=a+bTBF
(6)
式中a,b为常数。
1.3.3 最优游泳速度和能量消耗利用率
鱼类在运动过程中,采取不同的运动形式前进,为了使自身能量利用率最优化,通常使用最优游泳速度迁移。计算公式[19]如下:
(7)
式中a,b,c为拟合关系式中对应的常数。
能量消耗利用率(Rsc)是在一定游泳速度下比较自发性游泳和强迫式游泳下的能量消耗比。计算公式[19]如下:
(8)
1.3.4 自发性运动模式下各因素变量对耗氧率影响模型
自发性运动模式下,为了更好地量化分析鱼类在自发性运动下的呼吸代谢关系,该研究利用SPSS 19.0软件采用逐步回归,更好地表示多变量因素对耗氧率的影响,并可以反映出自发性运动下各因素对实验鱼耗氧率的贡献率。根据Steinhausen等[19]研究构建的多元线性回归模型如下:
logMo2=a+blogU+clogθ+dlogΔU
(9)
式中a,b,c,d为常数。
1.4 数据分析
实验数据采用SPSS 19.0进行统计,OriginPro 9.0进行作图,统计值描述用平均值±标准差 (mean±SD),差异显著水平为P<0.05。
2 结果与分析
2.1 自发性运动模式下游泳规律曲线
自发运动下各时间段的游泳速度U和耗氧率Mo2情况见图1。可知,在8 h的自发运动期间,鲢幼鱼的游泳速度U保持在0.08~0.5 BL·s-1的范围内,鲢幼鱼最大游泳速度达到0.5 BL·s-1,游泳规律为周期性运动;耗氧率Mo2变化规律与游泳速度U的变化规律几乎一致。
![]()
图1 自发性运动下耗氧率与游泳速度随时间变化曲线Fig.1 Curve of oxygen consumption rate and swimming speed in different time during spontaneous pattern
2.2 两种运动模式下游泳速度和耗氧率关系
在自发运动下,经比较发现鲢幼鱼的游泳速度与耗氧率关系(图2-a)参照方程(4)拟合方程为:Mo2=301.5+1 619U1.47(R2=0.64,P<0.05),该研究下,鲢幼鱼平均游泳速度达到最大值0.5 BL·s-1时,耗氧率为(866.45±18.90)mg·(kg·h)-1。该研究下标准代谢率为(305.24±12.30)mg·(kg·h)-1。
强迫式运动下,(25±1) ℃,体长为(11.60±0.42)cm鲢绝对临界游泳速度为(87.73±3.12) cm/s,相对临界游泳速度为(6.89±0.21) BL·s-1。耗氧率与游泳速度关系见图2-b,参照方程(4),拟合方程为Mo2=352+30.31U1.76(R2=0.94,P<0.05),该研究下标准呼吸代谢率为(355.16±22.30)mg·(kg·h)-1,游泳速度为0.5 BL·s-1时,实测耗氧率为(368.53±32.74)mg·(kg·h)-1,远远低于自发性运动该游泳速度下的耗氧率(866.45±18.90)mg·(kg·h)-1。
![]()
图2 自发性运动(a)和强迫式运动(b)模式下游泳速度与耗氧率关系Fig.2 Relationship between oxygen consumption rate and swimming speed in spontaneous pattern (a)and forced pattern (b)
2.3 自发性运动模式下加速度和旋转角度与耗氧率的关系
实验鱼在自发性运动期间,通过减速或加速和旋转来改变自身游泳的运动状态,类似的生理现象会产生呼吸代谢,加速度和旋转角度与耗氧率关系见图3,参照方程(4),拟合方程分别为:Mo2=278.35+61.66 ΔU1.33(R2=0.47,P<0.05);Mo2=337.6+0.5θ1.51(R2=0.33,P<0.05)。分析可见,在两种运动模式下耗氧率随加速度和旋转角度增加而呈增加的趋势。
![]()
图3 自发性运动下加速度(a)和旋转角度(b)分别与耗氧率关系Fig.3 Relationship between acceleration (a)and turning rate (b)with oxygen consumption rate in spontaneous pattern
2.4 两种运动模式下鲢的游泳行为
鱼类的运动主要靠摆尾进行(TBF),通过改变摆尾频率和摆尾幅度来改变游泳速度,进而完成自身运动。根据LoliTrack软件分析得出两种运动模式下摆尾频率TBF和游泳速度的关系(图4)以及摆尾频率TBF和耗氧率Mo2的关系(图5),参照方程(5)(6),自发性运动模式下:TBF=1.30+1.75U(R2=0.90,P<0.05)(图4-a);lnMo2=4.04+1.24x(R2=0.57,P<0.05)(图5-a)。强迫式运动模式下:TBF=1.60+1.71U(R2=0.94,P<0.05)(图4-b);lnMo2=5.5+0.25x(R2=0.96,P<0.05)(图5-b)。
![]()
图4 自发性运动(a)和强迫式运动(b)模式下游泳速度与摆尾频率关系Fig.4 Relationship between swimming speed and tail beat frequency in spontaneous pattern (a)and forced pattern (b)
![]()
图5 自发性运动(a)和强迫式运动(b)模式下摆尾频率与耗氧率关系Fig.5 Relationship between tail beat frequency and the oxygen consumption rate in spontaneous pattern (a)and forced pattern (b)
2.5 自发性运动模式下各因素变量对耗氧率影响模型
根据多元线性回归模型(9),模型中选取相关性R2最大的回归模型,回归模型主要参数见表1。
![]()
表1 多元线性回归模型及各影响因素贡献率Tab.1 Multiple linear regression model and contribution rate of each influencing factor
2.6 最优游泳速度和呼吸代谢率
根据公式(7)计算得出自发性运动和强迫式运动的最优游泳速度分别为0.4 BL·s-1和4.31 BL·s-1。不同运动模式下鲢幼鱼运动呼吸代谢情况见表2。
2.7 能量消耗利用率Rsc
该实验过程中,两种运动模式只存在唯一相同的相对游泳速度(0.5 BL·s-1)。根据公式(8)计算出该游泳速度下,鲢幼鱼的能量消耗利用率为
Rsc(U=0.5BL·s-1)=2.35
3 讨论
3.1 自发性运动模式下多因素与耗氧率关系分析
自发性运动模式下,鱼类的呼吸代谢受多种因素的影响,通过游泳速度对耗氧率的影响进行关系拟合相关性(R2=0.64)远远低于强迫式运动下游泳速度和耗氧率的相关性(R2=0.94),同样也低于房敏等[20]在25 ℃下鲢的研究成果(R2=0.997),造成这一现象的原因是每5 min计为一个计算单元,不能反映确切情况,导致数据分散度较高,降低了数据的精确性。该研究从拟合方程相关性分析,发现游泳速度与耗氧率的相关性(R2=0.64)远远高于加速度(R2=0.47)、旋转角度(R2=0.33)和摆尾频率(R2=0.57);从多变量回归模型相关性分析,游泳速度提供的贡献率(0.54)同样高于其它变量,但其它变量对鱼类的呼吸代谢同样提供一定的影响。对比游泳速度和耗氧率关系的拟合方程(R2=0.64)与多变量回归模型(R2=0.68)发现,二者的相关性相差并不大,所以在研究过程中并没有直接偏向于回归模型,只是为分析鱼类耗氧率多提供一种能量模型。
![]()
表2 两种运动模式下耗氧率情况Tab.2 Oxygen consumption rate in two different activity pattern
3.2 两种运动模式下的游泳行为分析
建立摆尾频率与游泳速度和耗氧率之间的关系对于鱼类游泳行为研究具有重要意义。自发性运动下鲢幼鱼的最大游泳速度只达到6.2 cm/s,且通过对游泳行为观察,实验鱼的转弯次数较多,摆尾频率变化较小,保持在1.3~2.2 s-1的范围内,摆尾幅度不明显,而且游泳方式和游泳位置相对稳定,游泳速度与时间成规律性变化,柯森繁等[21]对鲢在低流速下运动研究结果一致,游泳方式多采用“前进—滑行”,与蔡露等[22]对鳙鱼在低流速下的相似。
强迫式运动下,鱼类的摆尾频率随游泳速度的增加呈线性增加,与李会锋等[18]和蔡露等[22]对鱼类研究发现摆尾频率与游泳速度呈正相关结论一致。水流流速在5.5~41.5 cm·s-1范围内,实验鱼游泳方式相对稳定,运动范围主要位于实验区中下游,水流流速超过41.5 cm·s-1时,发现实验鱼为克服水流流速,摆尾振幅明显增加,运动位置逐渐转移到上游,且运动情况越来越不稳定。当游泳速度接近临界游泳速度,实验鱼多次交替使用加速游动和滑行的方式,这一现象同蔡露等[10]和刘慧杰等[16]对鳙鱼游泳行为研究一致,说明鱼类在一定范围内通过改变自身运动方式来应对外界因素的干扰,进而合理利用自身能量。在鱼道建设过程中,过鱼对象在什么样的流速范围内,采取什么样的游泳行为,以及能达到的游泳速度是多少,鱼类运动过程中的喜好流速,都将对鱼类上溯是否成功起着决定性的因素。
3.3 两种运动模式下呼吸代谢研究比较
鱼类游泳速度与耗氧率变化密切相关。两种运动模式下,鲢幼鱼在自发性运动下的标准代谢率略低于强迫式运动,但两者差异性不显著(P>0.05)。游泳速度的幂指数c代表鱼类在该运动情况下的游泳能力[23]和呼吸代谢指数[24],通过文献查阅鱼类的该指数范围在1.1~3.0,该研究的结果分别是1.47和1.76。经比较分析,鲢幼鱼在强迫式运动下的游泳能力优于自发性运动,说明在一定的流速范围内,游泳能力会随流速的增加而提高,强迫式运动更能激发鱼类的游泳潜力;但自发性运动呼吸代谢能量利用率要优于强迫式运动,表明自发性运动下鱼类根据自身生理需求来进行能量利用,更好进行能量分配。
在两种运动模式唯一相同的相对游泳速度为0.5 BL·s-1下,能量消耗利用率为2.35,说明在该平均速度下,与强迫游泳相比,自发性游泳运动期间使用将近2.4倍的能量来完成自身的运动。这与Steinhausen等[19]的理论计算相似,表明自发性运动造成较大的能量消耗。经过对文献的分析发现能量消耗利用率的研究范围为1.4~22,且能量消耗利用率的值并不固定,造成这一现象的主要原因是在实验过程中温度的差异、实验鱼的生长状况以及实验装置的影响等。对于该研究中能量消耗利用率的大小是否能表明能量利用情况的优劣,该内容并没找到相应的结论分析,还需进一步的研究进行论证分析。
两种运动模式下的最优游泳速度比较分析,发现强迫式运动下的最优游泳速度(4.31 BL·s-1)要远远高于自发性运动(0.4 BL·s-1),差异性显著(P<0.05)。自发性运动下的游泳速度要远远低于Blaxter[25]研究中建议作为多数鱼类最优巡航的相对速度(2 BL·s-1),造成该现象的原因与实验装置的大小限制了自发性运动下的游泳速度大小相同,实验鱼的游泳行为造成一定的影响,所以在该运动模式下的最优速度会同样受到影响。强迫式运动的最优游泳速度高于2 BL·s-1,文献[26]表明鱼类的最优巡航速度与体长的0.4次方成正比,而刘慧杰等[27]对鱼类体长和相对临界游泳速度关系发现,相对临界游泳速度随体长的增加而降低,造成此种现象的原因尚不明确,所以对实验鱼最优游泳速度的影响因素还有待进一步研究。Webb[28]研究表明在强迫式运动模式下的最优游泳速度通常与正常状态下鱼类的最佳游泳速度差异性不显著(P>0.05),所以该运动模式下的最优游泳速度经常被作为鱼类行为学研究一个标准,对鱼类游泳能力和行为研究具有重要意义。
两种运动模式下的最优游泳速度相差很大,但耗氧率差异性不显著(P>0.05)。这说明在鱼类运动过程中最优游泳速度可能不是唯一表明鱼类最佳游泳状态的标准,而存在表征能量利用率最高的最优呼吸代谢耗氧率。鱼类的最优耗氧率表明鱼类此时的运动为全部的有氧运动来提供自身能量,因为一旦由无氧运动进行参与,便会增加氧负债,这对生物是不利的。蔡露等[29]对虹鳟研究发现当鱼类游泳速度达到70%~80% Ucrit时,便有部分的无氧呼吸参与。该研究中,当游泳速度达到70%Ucrit时,耗氧率为(848.97±17.68)mg·(kg·h)-1,最优游泳速度的耗氧率低于该速度的耗氧率,验证了本研究最优游泳速度下,鱼类呼吸方式为100%有氧呼吸。对于最优呼吸代谢耗氧率能否作为研究鱼类游泳特性的工具,还需在以后的研究中进行模拟自发性运动的实验证明。
综上,需要充分考虑鱼类在不同运动模式下的游泳特性,如设计鱼道时严格控制流速,以防鱼类过度消耗能量,导致疲劳进而造成过坝失败,具体数值还需要后续实验进一步研究。
[1]袁 喜,李丽萍,涂志英,等.鱼类生理和生态行为对河流生态因子响应研究进展[J].长江流域资源与环境,2012,21(S1):24-29.
[2]杨君兴,潘晓赋,陈小勇,等.中国淡水鱼类人工增殖放流现状[J].动物学研究,2013,34(04):267-280.
[3]诸葛亦斯,刘德富,谭红武.鱼类栖息地流速适宜性曲线实验方法[J].水利学报,2013,44(S1):1-7.
[4]曹庆磊,杨文俊,周良景.国内外过鱼设施研究综述[J].长江科学院院报,2010,27(05):39-43.
[5]王从锋,陈明明,刘德富,等.基于葛洲坝1号船闸模型的水流诱鱼试验研究[J].长江流域资源与环境,2016,25(06):974-980.
[6]郑金秀,韩德举.国外高坝过鱼设施概况及启示[J].水生态学杂志,2013,34(04):76-79.
[7]汤荆燕,高 策,陈 昊,等.不同流态对鱼道进口诱鱼效果影响的实验研究[J].红水河,2013,32(01):34-39,44.
[8]刘志雄,周 赤,黄明海.鱼道应用现状和研究进展[J].长江科学院院报,2010,27(4):28-31,35.
[9]涂志英,袁 喜,王从锋,等.亚成体巨须裂腹鱼游泳能力及活动代谢研究[J].水生生物学报,2012,36(4):682-688.
[10]蔡 露,房 敏,涂志英,等.与鱼类洄游相关的鱼类游泳特性研究进展[J].武汉大学学报(理学版),2013,59(4):363-368.
[11]Ohlberger J,Staaks G,Hölker F.Estimating the active metabolic rate (AMR)in fish based on tail beat frequency (TBF)and body mass[J].J Exp Zool A Ecol Genet Physiol,2007,307A(5):296-300.
[12]柯森繁,高 柱,刘国勇,等.基于Matlab的鱼类游泳动力学分析[J].水生生物学报,2016,40(5):985-991.
[13]熊 锋,王从锋,刘德富,等.松花江流域青鱼、草鱼、鲢及鳙突进游速比较研究[J].生态科学,2014,33(2):339-343.
[14]张永泉,尹家胜,徐革峰,等.水温和体重对白斑红点鲑临界游泳速度和游动耗氧率的影响[J].水生生物学报,2015,39 (4):661-668.
[15]王从锋,向经文,刘德富,等.一种利用船闸实现过鱼的装置及方法[P]:湖北,CN103485314A[P],2014-01-01.
[16]刘慧杰,王从锋,刘德富,等.不同运动状态下鳙幼鱼的游泳特性研究[J].南方水产科学,2017,13(2):85-92.
[17]Tu Z Y,Yuan X,Han J C,et al.Aerobic swimming performance of juvenileSchizothoraxchongi(Pisces,Cyprinidae)in the Yalong River,southwestern China[J].Hydrobiologia,2011,675(1):119-127.
[18]李会锋,蔡德所,杨培思.鲢幼鱼游泳能力及游泳行为的试验研究[J].水生态学杂志,2016,37(3):88-92.
[19]Steinhausen M F,Steffensen J F ,Andersen N G.The effects of swimming pattern on the energy use of gilthead seabream (Sparus aurata L.)[J].Mar Freshw Behav Phy,2010,43(4):227-241.
[20]房 敏,蔡 露,高 勇,等.温度对鲢幼鱼游泳能力及耗氧率的影响[J].水生态学杂志,2013,34(3):49-53.
[21]柯森繁,陈渴鑫,罗 佳,等.鲢顶流游泳速度与摆尾行为相关性分析[J].水产学报,2017,41(3):401-406.
[22]蔡 露,涂志英,袁 喜,等.鳙幼鱼游泳能力和游泳行为的研究与评价[J].长江流域资源与环境,2012,21(Z2):89-95.
[23]房 敏,蔡 露,高 勇,等.运动消耗对草鱼幼鱼游泳能力的影响[J].长江流域资源与环境,2014,23(6):816-820.
[24]Cai L,Liu G Y,Taupier R,et al.Effect of temperature on swimming performance of juvenile Schizothorax prenanti[J].Fish Physiol Biochem,2014,40(2):491-498.
[25]Blaxter J H S.Swimming speed of fish[C].Fish conference on fish behavior in relation to techniques and tactics,Bergen:1969.
[26]王 萍,桂福坤,吴常文.鱼类游泳速度分类方法的探讨[J].中国水产科学,2010,17(5):1137-1146.
[27]刘慧杰,王从锋,朱良康,等.鲢鳙幼鱼临界游泳速度的比较研究[J].水生态学杂志,2016,37(4):63-69.
[28]Webb P W.Composition and mechanics of routine swimming of rainbow trout,oncorhynchus mykiss[J].Can J Fish Aquat Sci,1991,48(4):583-590.
[29]Cai L,Chen L,Johnson D,et al.Integrating water flow,locomotor performance and respiration of Chinese sturgeon during multiple fatigue-recovery cycles[J].Plos One,2014,9(4):e94345.
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!



