
通讯:13983250545
微信:ycsh638
QQ:469764481邮箱:ycsh6318@163.com
论文:重庆养殖场鳜群体微卫星遗传多样性研究
重庆养殖场鳜群体微卫星遗传多样性研究
范士琦,冯婧昀,苗晓敏,郭慧,陶怡曦,李云
(1.西南大学水产学院,渔业与水产生物技术实验室,重庆 400715;2.西南大学淡水鱼类资源与生殖发育教育部重点实验室,重庆市水产科学重点实验室,重庆 400715)
鳜(Siniperca chuatsi)俗称桂花鱼、翘嘴鳜,隶属于硬骨鱼纲(Osteichthyes),鲈形目(Perciformes),科(Serranidae),鳜亚科(Siniperinae),鳜属(Siniperca),广泛分布于中国长江和黑龙江水系及其附属湖泊水库以及朝鲜半岛、越南部分地区,为名贵水产品[1-3]。2020 年我国鳜养殖的总产量达37 万t,广东生产的鳜苗种占全国的95%以上[3]。鳜作为重要养殖鱼类,国内学者对其形态特征、养殖性能、种质资源、生理机能、遗传育种等方面开展了广泛研究[4-8],取得了丰富的成果,目前已经报道的鳜新品种有“华康1 号”“长珠杂交鳜”“广清1号”“鼎鳜1 号”等[9-12]。尽管长江流域有着丰富的天然鳜种质资源,但位于长江上游的川渝地区鳜养殖产业起步较晚。近几年,重庆市鳜消费市场需求旺盛,鳜苗种需求持续增加,外购苗种质量不稳定,加上长途运输易产生应激反应,养殖风险大,制约了重庆鳜养殖业的发展[13]。
遗传多样性可以直接体现种群长期进化和发展中的遗传变异,因此开展鱼类遗传多样性的研究,可为了解鱼类的种质资源现状和遗传改良奠定基础[14-15]。微卫星标记技术是目前发展最快的一类分子标记,在鱼类遗传发育、DNA 指纹图谱制作分析、遗传连锁图谱构建和标记辅助选择中广泛使用[8]。微卫星DNA 在真核及原核生物基因组中被普遍应用,其特点相较于其他诸如RFLP、RAPD、AFLP 分子标记技术而言,多态性更高[16-17]。现通过微卫星DNA 遗传多样性,对重庆地区3 个养殖场的鳜群体的遗传多样性和遗传结构进行分析评价,拟为重庆鳜养殖种质资源调查提供基础数据,为优质苗种生产提供参考。
1 材料与方法
1.1 材料
活体鳜采自重庆市潼南区兴水养殖场(XS)、名优鱼养殖场(MY),以及重庆市巫溪县鳜养殖场(WX)。3 个群体样本的外部形态特征和可数性状标准均符合《鳜》(SC 1038—2000)的分类特征。各养殖场均随机采集样本50 尾,分别测量体高和体长。其中,MY 养殖场鳜平均体质量为(73.76±20.16)g,平均体长为(14.21±1.09)cm;XS 养殖场鳜平均体质量为(49.60±14.91)g,平均体长为(12.79±1.50)cm;WX 养殖场鳜平均体质量为(32.94±7.25)g,平均体长为(11.03±0.77)cm。捕捞后立即取尾鳍固定在95%乙醇中,于-80 ℃冰箱保存备用。
1.2 微卫星DNA 标记
所有鳜样本基因组DNA 提取均采用酚-氯仿抽提法[18]。根据文献[19-21],筛选出23 对鳜微卫星引物(由上海生工合成)。从3 个鳜群体中分别任选2 尾鳜的模板DNA 与23 对引物进行PCR 扩增筛选出10 对扩增效率高、多态性较好的引物(表1),并合成荧光基团标记引物(上海生工)。
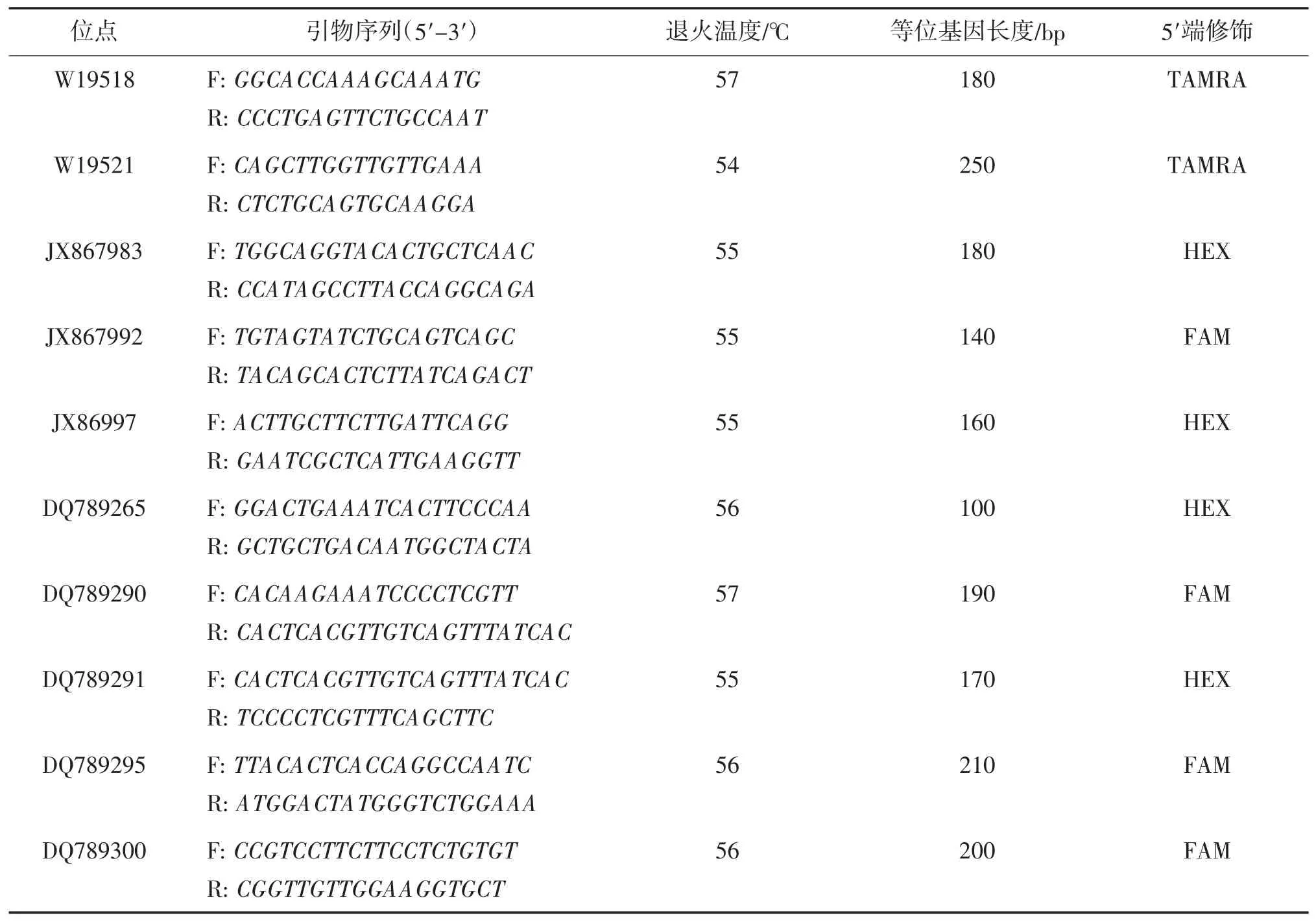
表1 10 对荧光引物基本信息
其中,微卫星引物JX867997、JX867983、JX867992参考文献[20],另外7 个引物参考文献[19,21]。10对引物的正向引物(5′端)添加HEX(蓝色)或FAM(绿色)或TAMRA(黑色)荧光标记,反向(3′端)不添加,然后分别与模板DNA 进行PCR 扩增反应。并通过ABI3730XL 遗传分析系统对产物进行STR 分型。
1.3 数据处理
采用Genemapper 软件,分析SSR 数据,统计分析包括等位基因数(Na)、有效等位基因数(Ne)、观测杂合度(Ho)、期望杂合度(He)、香农威纳指数(H′)、基因流(Nm)、基因分化系数(FST)等参数;哈迪温伯格平衡(Hardy-Weinberg)检验遗传距离和遗传相似系数;使用PIC_Calc 0.6 软件计算多态信息含量(PIC)。
2 结果与分析
2.1 群体遗传多样性
用10 个多态性好的微卫星引物共检测到95个等位基因(表2),Na为4~22,均值为9.5。就PIC而言,DQ789300 的多态性为中度(0.25<PIC<0.5)外,另外9 个多态性位点均有高度多态性(PIC>0.5)。10 个微卫星位点FIS值为-0.390 9~0.146 9,FIT值为-0.339 4~0.188 6,平均值为-0.051 8;FST值为0.012 2~0.087 6。就Nm而言,除了DQ789391 的值<4 外,其微卫星位点的Nm值均>4。
![]()
表2 10 个微卫星位点的固定指数(F)和Nm①
筛选出的10 个微卫星位点在3 个鳜群体中的遗传多样性参数见表3。由表3 可见,鳜群体的Na均值为6.4~7.4,Ne均值为3.635~3.767,Ho均值为0.733~0.755,各群体He均值为0.674~0.711。3 个鳜群体I 均值为1.378~1.427,其中XS 群体最低,MY群体最高。
![]()
表3 3 个鳜群体微卫星遗传多样性参数①
2.2 群体遗传结构
3 个鳜群体间遗传相似系数(对角线上)和遗传距离(对角线下)见表4。由表4 可见,不同鳜群体间的遗传距离值为0.087 1~0.136 0,遗传相似系数值为0.889 5~0.916 6,>0.05,其中MY 群体与XS 群体间相似系数值(0.916 6)高于MY 群体与WX 群体间遗传相似系数(0.872 8)。对重庆地区3 个鳜群体的分子方差分析结果表明(表5),95.44%的遗传变异来自个体内,4.47%的变异来自群体间,群体内变异组成为0.09%。
![]()
表4 3 个鳜群体间遗传相似系数(对角线上)和遗传距离(对角线下)
![]()
表5 3 个鳜群体的分子方差分析(AMOVA)
3 讨论
3.1 3 个鳜群体的遗传多样性
微卫星标记分析遗传多样性会受到种群样本量的影响,通常认为用于微卫星标记研究的样本量在≥20 个时,才能稳定反映出等位基因和有效基因数[22]。本试验的各群体的样本量为50 尾,可以反映出3 个鳜群体间的遗传变异程度。Buchanan 等[23]研究发现,高度多态表现为PIC>0.5,中度多态时PIC值为0.25~0.50,而PIC<0.5 时,则为低度多态;另外,微卫星位点选择标准是等位基因数在4 个以上[24]。本研究中,除了DQ789300 位点属于中度多态外,其余9 个微卫星位点的PIC 均在0.5 以上,属于高度多态;从Na来看,除DQ789300 外,其余微卫星位点的Na均≥4,能将群体遗传变异程度很好表现出来。
本研究所使用的微卫星位点所获得的遗传分析结果是可靠的,引物JX867983、JX867992 和JX86997,在150 个样本中观察到的等位基因数为7~11 个,低于文献[25]运用相同引物对3 种鳜属鱼类进行遗传分析所得到的等位基因数(11~19 个),这可能与本研究样本为同种样本有关。
本研究中,MY、XS、WX 群体的Na和Ne平均值分别为6.4,7.4,6.9 和3.675,3.635,3.767,不同群体之间差异较小。Na平均值与李珊[26]对清水江野生斑鳜群体的研究结果相差不大(Na=3.668 0)。Takezaki等[27]通过微卫星分析得出,群体间有遗传多样性表现在杂合度为0.3~0.8。MY、XS、WX 群体的平均Ho和He分别为0.733、0.755、0.736 和0.711、0.682、0.674,杂合度较高,群体变异丰富[18]。彭敏燕等[28]在对3 个鳜野生群体的微卫星遗传多样性研究中得出平均Ho和He分别为0.255 4~0.698 9 和0.710 0~0.744 6,低于本研究中群体结果平均Ho,稍高于本研究群体平均He值。从H′来看,各群体均值为1.378~1.427,高于梅秋兰等[29]对鳜原种群体和鳜群体使用微卫星技术研究得出的H′均值(1.078 和0.689)。本研究中,重庆市3 个鳜群体遗传多样性较高,提示3 个鳜养殖群体苗种来源的亲代繁育群体数量较大,遗传异质性较高,也反映出苗种产地人工选择压力小等原因[30]。
3.2 3 个鳜群体间的遗传结构
固定指数是研究遗传分化的重要指标,它包括FIT、FIS和FST3 种参数[31-32]。其中,FIS表示群体偏离随机交配的程度,当FIS<0 认为群体在该位点存在杂合子过剩现象,FIS>0,则表示杂合子缺失,FIS=0 时,即为达到平衡[33]。在本研究中,除了W19518、DQ789290、DQ789391 位点FIS>0 外,其他位点的FIS均<0,表明存在杂合子过剩,异交程度高。群体间的遗传分化程度可以通过FST揭示出来,分化程度较小时FST<0.05,分化程度中等时为0.05~0.15,当FST>0.15 时,表示群体间的分化程度高[34]。本研究中,有70%的微卫星位点的FST值均<0.05,表明3 个群体间的分化程度低。针对Nm而言,一般认为如果Nm>1,则表示不会有显著的遗传分化,当Nm>4 时,则被认为分化程度很小[35],本研究中,各位点的Nm均>4,也证明3 个鳜群体间均未出现遗传分化现象。而根据AMOVA 来看,个体内部变异(95.44%)是遗传变异的最主要源头,群体间的变异程度仅占4.47%,这也符合FST得出的分析结果。
研究群体多样性的重要因素之一是遗传距离的大小,其与群体分化时间成正相关[36]。根据3 个鳜鱼群体间的遗传相似度、遗传距离数据,MY 群体与XS 群体遗传相似度最大(0.916 6),且遗传距离最短,而WX 群体与其他2 个群体的遗传相似度略小,遗传距离略大。文献[37]表明,同种间的群体遗传相似度为0.80~0.97,遗传距离为0.03~0.20。本研究结果显示,遗传相似度为0.872 8~0.916 6,遗传距离为0.087 1~0.136 0,可判定3 个群体为同一物种,且群体分化时间短。根据调查,XS、WX2 个群体苗种分别来源于广东清远、佛山,MY 群体属于重庆本地储养亲体的自繁苗种,其亲体从广东佛山引入,提示3 个群体的亲本可能来源于共同的祖代群体,也与广东鳜(翘嘴鳜)的养殖祖代群体于1980 年代引自长江中下游的历史相吻合[1,3]。
许多研究发现,养殖群体比野生群体发生瓶颈效应和种质同质化的可能性更大,这些都会导致遗传基因的丢失和群体遗传性的降低。在对来自湘江的野鲤养殖群体和野生群体的研究中发现,2 个群体间的遗传变异水平很大[38];赵祥等[39]研究后发现,对比野生群体来看,黄姑鱼养殖群体的遗传多样性降低十分显著。在对鳜的遗传多样性研究中,方展强等[40]研究得出鳜养殖群体与野生群体之间遗传差异十分显著,养殖群体要显著低于野生群体;梅秋兰等[30]也基于微卫星对鳜养殖群体和原种群体的遗传差异进行了分析比较,得出鳜原种群体遗传多样性较为丰富的结论。因此,在原种场亲本群体和后备群体的选择时,选择遗传距离相对较远的亲本,并开展群体遗传多样性分析,对保持亲本群体的遗传多样性非常有必要性[41]。
4 结语
本研究运用微卫星标记对重庆3 个鳜养殖群体进行分析,结果表明,3 个鳜群体内的遗传多样性较高,群体间的遗传分化水平低,表明3 个群体内保留有较高的遗传异质性,群体间有较近的亲缘关系,反映了群体亲本源自共同的祖代。在生产中,要提高苗种质量、存活率和养殖性能,需加强亲本选育过程的遗传选择压力,提高已育成良种的生产力和覆盖率。
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!



